尋媶戣栚
- 乽泱桿摫尰徾偵偍偗傞丄嵶朎奜僔僌僫儖偲嵶朎撪場巕偺揮幨場巕傪夘偟偨摑崌儊僇僯僘儉乿
尰嵼傑偱偺尋媶偲惉壥
- 丂戝妛堾廋巑壽掱傑偱偼丄柍惈惗怋傪峴偆儈僒僉儅儊僀僞儃儎偺埻橐峯忋旂嵶朎傪梡偄偨尋媶傪峴偄丄弌夎偍傛傃嵞惗夁掱偵偍偗傞暘壔懡擻惈偺惂屼偺偟偔傒傪夝愅偟偨丅傑偢丄埻橐峯忋旂嵶朎桼棃偺攟梴嵶朎偵懳偟偰嵶朎憹怋梷惂偍傛傃嬅廤妶惈傪帩偮場巕傪惛惢偟偨丅偙偺傛偆側妶惈偼偙偺埻橐峯忋旂攟梴嵶朎偑扙暘壔偟偝傜偵庬乆偺慻怐傊偲嵞暘壔偡傞偺傪梷偊丄埻橐峯忋旂偺暘壔忬懺偺堐帩偵娭梌偟偰偄傞丅偙偺場巕傪丄嵟廔揑偵俀偮偺儁僾僠僪傪娷傓夋暘偵傑偱惛惢偟丄傾儈僲巁攝楍傪抐曅揑偵僔乕僋僄儞僗偟偰丄偦偺攝楍傪傕偲偵慡挿cDNA傪僋儘乕僯儞僌偟偨丅俀偮偺儁僾僠僪偼C宆儗僋僠儞(TC14-1)偲椶帡惈偑崅偔丄曅曽偼埲慜偵cDNA抐曅偑曬崘偝傟偰偄偨儗僋僠儞乮TC14-2乯偱偁傝丄曅曽偼怴婯儗僋僠儞偱偁偭偨乮TC14-3偲柦柤乯丅師偵丄戝挵嬠傪梡偄偰丄TC14-2,3偺儕僐儞價僫儞僩僞儞僷僋幙傪偮偔傝丄摐嵔寢崌擻傪挷傋偨丅偡傞偲丄偙傟傜偺儗僋僠儞偼僇儖僔僂儉埶懚揑偵僈儔僋僩乕僗偵寢崌偟偨丅偟偐偟丄攟梴嵶朎偵懳偟偰偼俁庬椶偺偆偪TC14-3偺傒偑嵶朎憹怋梷惂偍傛傃嬅廤妶惈傪帩偭偰偄偨丅傑偨丄妋偐偵TC14-3偼埻橐峯忋旂傊偺暘壔傪桿摫偟丄堐帩偡傞嶌梡傪帩偭偰偄傞偙偲偑妋擣偝傟偨丅攟梴嵶朎偵懳偡傞TC14-3偺妶惈偼僈儔僋僩乕僗偺揧壛偱姰慡偵幐傢傟偨偺偱丄僈儔僋僩乕僗寢崌擻偑偙傟傜偺妶惈偵昁梫偱偁傞偙偲偑傢偐偭偨丅夁嫀偺抦尒側偳傕娷傔偰峫偊傞偲丄TC14-3偼宍懺宍惉椞堟偵埻橐峯忋旂嵶朎傪嫙媼偟丄宍懺宍惉傪庤彆偗偟偰偄傞偙偲偑帵嵈偝傟偨(Development,
2001)丅
丂攷巑壽掱偐傜尰嵼傑偱偼丄儅儃儎偲儐僂儗僀儃儎傪梡偄偰丄扨弮側恄宱宯傪帩偮儂儎梒惗偺恄宱敪惗偵偍偗傞揮幨挷愡儊僇僯僘儉傪尋媶偟偨丅儅儃儎-僔僫僾僩僞僌儈儞(Hr-syt)偼丄僔僫僾僗揱払偵娭傢傞暘巕偱丄梒惗婜偐傜偡傋偰偺僯儏乕儘儞偱敪尰偡傞丅偦偺敪尰偼Hr-syt偺5乫忋棳3.4kbp偺僾儘儌乕僞乕椞堟偱惂屼偝傟傞偙偲偑傢偐偭偰偄偨丅偦偙偱丄僨儕乕僔儑儞僐儞僗僩儔僋僩偵傛傞儗億乕僞乕偺敪尰夝愅傪峴偄埲壓偺偙偲傪柧傜偐偵偟偨丅
丂僯儏乕儘儞偱偺敪尰偵娭傢傞椞堟偼暋悢懚嵼偟偨(Dev.
Biol.,
2002)丅偦傟傜偺偆偪偺俁偮傪200bp埲壓偺椞堟偵峣傝崬傫偩丅侾偮栚偼-558偐傜-477偺椞堟偱丄偙偺椞堟傪寚偔偲堎強揑側儗億乕僞乕敪尰偑傒傜傟傞帠偐傜丄堎強揑敪尰傪梷惂偡傞偙偲偱敪尰傪僯儏乕儘儞偵尷掕偡傞偨傔偺椞堟偱偁傞帠偑帵嵈偝傟偨丅俀偮栚偼-2900偐傜-2700偺椞堟偱丄偙偺椞堟傪寚偔偲僾儘儌乕僞乕妶惈偑庛偔側傝丄僞儞僨儉偵俀偮暲傋傞偲僾儘儌乕僞乕妶惈偑嫮偔側偭偨偙偲偐傜丄僗僥僢僾傾僢僾僼傽僋僞乕寢崌僒僀僩偱偁傞偲梊憐偝傟偨丅俁偮栚偼5乫UTR忋偺+100偐傜+200偺椞堟偱丄偙偺椞堟傪寚偔偲僯儏乕儘儞偱偺敪尰偑側偔側偭偨丅偙偺椞堟偵偼E-box俀偮傪娷傓摿挜揑側攝楍偑暲傫偱偄傞偙偲偐傜丄偦偙偵helix-loop-helix僞僀僾偺揮幨場巕偑嶌梡偟丄僯儏乕儘儞偱偺敪尰傪妶惈壔偟偰偄傞偙偲偑梊憐偝傟偨丅尰嵼丄偙偺椞堟偵嶌梡偡傞揮幨場巕傪挷傋偰偄傞丅
丂儅儃儎syt僾儘儌乕僞乕傪儐僂儗僀儃儎偵摫擖偟偨偲偙傠丄僯儏乕儘儞摿堎揑側敪尰偑桿摫偝傟偨丅媡偵丄儐僂儗僀儃儎syt僾儘儌乕僞乕傕儅儃儎偲儐僂儗僀儃儎偺椉庬偵偍偄偰僯儏乕儘儞偱偺敪尰傪堷偒婲偙偟偨丅偙偺帠偐傜丄俀庬娫偵嫟捠偺僯儏乕儘儞摿堎揑側敪尰惂屼椞堟偑懚嵼偡傞帠偑梊憐偝傟偨丅偟偐偟丄扨弮側僾儘儌乕僞乕攝楍斾妑偱偼椶帡攝楍傪尒弌偡帠偼偱偒偢丄尰嵼丄椉庬偱嫟捠偵摥偔僯儏乕儘儞摿堎揑側敪尰惂屼椞堟傪扵嵏拞偱偁傞丅偪側傒偵丄儅儃儎偱摨掕偟偨僯儏乕儘儞摿堎揑側敪尰惂屼椞堟乮-558偐傜-477偍傛傃-2900偐傜-2700乯偼儐僂儗僀儃儎偱傕摥偔帠偑傢偐偭偨丅
尋媶栚揑
-
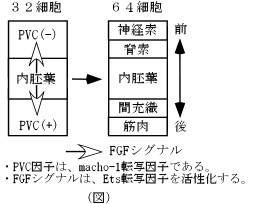
巹偼丄惗柦偑侾偮偺棏偐傜恊屄懱傑偱宍偯偔傜傟傞巇慻傒偵嫽枴傪帩偭偰偍傝丄偦傟傪夝柧偟偰偄偔偙偲偱恖椶偵偲偭偰桳塿側抦揑嵿嶻偑摼傜傟傞偲峫偊偰偄傞丅嬤擭丄愐嶕摦暔偺儂儎傪梡偄偨尋媶偑悽奅拞偱拲栚偝傟偰偒偰偍傝丄僎僲儉夝愅傗EST夝愅側偳傕峴傢傟幚尡摦暔偲偟偰偺壙抣偑崅傑偭偰偄傞丅傑偨丄儂儎偼僔儞僾儖偱幚尡偑慻傒傗偡偄丅幚嵺丄惣揷傜偵傛偭偰儂儎泱敪惗夁掱偵偍偗傞拞泱梩偺僷僞乕僯儞僌偑柧傜偐偵偝傟偮偮偁傝丄偦偺儌僨儖偼旕忢偵僔儞僾儖側傕偺偱偁傞乮恾乯丅棏嵶朎幙撪偱屻曽偵埵抲偡傞儅僀僆僾儔僘儉(PVC場巕)偺懚嵼壓偱偼娫廩怐傗嬝擏側偳偺屻曽慻怐丄PVC場巕旕懚嵼壓偱偼愐嶕丄恄宱嶕側偳偺慜曽慻怐偑嶌傜傟傞丅堦曽偱俁俀嵶朎婜偐傜俇係嵶朎婜偺娫偵撪泱梩偐傜偺FGF僔僌僫儖偵傛傞桿摫傪庴偗傞偲愐嶕傗娫廩怐偑桿摫偝傟丄媡偵FGF僔僌僫儖偺桿摫傪庴偗側偄偲嵶朎偼僨僼僅儖僩偺塣柦偱偁傞恄宱嶕傗嬝擏偵暘壔偡傞丅偝傜偵丄PVC場巕偺幚懱偼僕儞僋僼傿儞僈乕僪儊僀儞傪帩偪嬝擏偺僨僞乕儈僫儞僩偟偰摨掕偝傟偨macho-1偱偁傞丅偡側傢偪丄macho-1傪帩偭偨嵶朎偑FGF僔僌僫儖傪庴偗庢傞偲娫廩怐偑桿摫偝傟丄帩偨側偄嵶朎偑僔僌僫儖傪庴偗庢傞偲愐嶕偑桿摫偝傟傞丅桿摫傪攠夘偡傞FGF僔僌僫儕儞僌宱楬偵偍偄偰偼丄懠偺懡偔偺惗暔摨條偵FGFR丄Ras丄Raf丄MEK丄MAPK丄Ets偺儕儞巁壔僇僗働乕僪偑曐懚偝傟偰偄傞丅偟偐偟丄偙傟傜偺場巕偑偳偺傛偆偵暘巕揑偵憡屳嶌梡偟偰埲崀偺堚揱巕敪尰傂偄偰偼拞泱梩偺僷僞乕儞僯儞僌偵娭傢偭偰偄傞偐偼尰嵼傢偐偭偰偄側偄丅杮尋媶偱偼丄偙傟傜偺帠傪傆傑偊偰FGF僔僌僫儖偺壓棳場巕Ets偺揮幨挷愡婡峔傪拞怱偵偟偰丄儂儎泱偵偍偗傞拞泱梩偺僷僞乕僯儞僌婡峔傪峏偵怺偔夝愅偡傞丅摿偵丄macho-1偲Ets偲偑偳偺傛偆側巇慻傒偱嫤挷揑偵摥偒嵶朎偺塣柦寛掕傪挷愡偟偰偄傞偺偐夝愅偟偨偄丅
尋媶撪梕
- 丂泱桿摫偵偼嵶朎奜偐傜棃傞僔僌僫儖偲嵶朎撪偺場巕偑娭學偟偰偄傞丅偙偺擇偮偺梫慺偑丄桿摫偑婲偙傞嵶朎撪偱摑崌偝傟傞偙偲偵傛傝丄嵶朎偺塣柦側偳偑寛掕偝傟傞丅嵶朎撪偺場巕偼丄桿摫僔僌僫儖偑棃偨偲偒偺墳摎嵶朎偺墳摎擻傕偟偔偼斀墳擻偺惂屼偵娭傢偭偰偄傞丅偙偙偱夝愅偟傛偆偲偟偰偄傞宯偱偼丄嵶朎奜僔僌僫儖偼FGF偱偁傝丄嵟廔揑偵Ets揮幨場巕偺妶惈壔傪堷偒婲偙偡丅斀墳擻傪寛傔偰偄傞嵶朎撪揮幨場巕偼macho-1偱偁傞丅偙偺擇偮偺揮幨場巕偑摥偔慻傒崌傢偣偵傛傝丄4庬椶偺慻怐乮恄宱嶕丒愐嶕丒娫廩怐丒嬝擏乯偑儂儎泱偺愒摴椞堟偵宍惉偝傟傞丅偟偐偟丄偳偺條側暘巕婡峔偱丄撪嵼場巕(macho-1)偲僔僌僫儖揱払宯(Ets)偺忣曬偑嵶朎撪偱摑崌偝傟傞偐偼晄柧偱偁傞丅偙偺宯偼偡偱偵僉乕暘巕偑傢偐偭偰偄傞偺偱丄桿摫尰徾傪夝愅偡傞忋偱儌僨儖働乕僗偲側傝偆傞丅幚嵺偵偼偄傠偄傠側働乕僗傪憐掕偟偮偮丄夝愅傪埲壓偺傛偆偵峴偭偰偄偔丅
丂Ets偵偼偄傠偄傠側場巕偑嫤挷揑偵摥偄偰偄傞偲偄偆曬崘偑偁傞丅偦偙偱丄Ets偲macho-1偑捈愙寢崌偟偰嶌梡偟偰偄傞偐傪丄偦傟偧傟偺儕僐儞價僫儞僩僞儞僷僋幙傪戝挵嬠偵嶌傜偣偰挷傋傞丅庤朄偲偟偰偼僾儖僟僂儞傾僢僙僀朄傕偟偔偼峈懱捑崀朄傪梡偄傞丅傑偨丄Ets偵寢崌偟偆傞懠偺場巕偵偮偄偰偺扵嵏傕峴偆丅
丂師偵丄Ets僞儞僷僋幙偺嬊嵼傪挷傋傞偨傔偵丄Ets僞儞僷僋幙傪摿堎揑偵擣幆偡傞峈懱偺嶌惉傪峴偆丅嶌惉偵巊偆峈尨偼儕僐儞價僫儞僩Ets僞儞僷僋幙傪梡偄傞丅偱偒傟偽儕儞巁壔偟偨Ets偲儕儞巁壔偟偰偄側偄Ets傪擣幆偡傞峈懱傪偦傟偧傟嶌惉偱偒傟偽FGF僔僌僫儖宱楬偺摥偔夁掱傪壜帇壔偡傞偙偲偑偱偒傞偩傠偆丅
丂尰嵼傑偱偺抦尒偱偼丄FGF僔僌僫儕儞僌偵埶懚偟偰愐嶕嵶朎偱偼Brachyury偺堚揱巕敪尰偑64嵶朎婜偵妶惈壔偝傟傞丅偙傟偵懳偟丄娫廩怐嵶朎偱偼丄64嵶朎婜偵嬝擏傾僋僠儞偺堚揱巕敪尰偑偄偒側傝梷惂偝傟傞丅偙偺擇偮偺尰徾偵偼Ets揮幨場巕偑昁梫偱偁傞丅偦偙偱丄Ets偑壓棳堚揱巕傪偳偺傛偆偵惂屼偡傞偐傪挷傋傞偨傔偵丄Ets偲揮幨妶惈壔僪儊僀儞(VP16)傕偟偔偼梷惂壔僪儊僀儞(EnR)偲傪梈崌偝偣偨倣RNA傪嶌傝丄儂儎棏偵尠旝拲擖偟偰挷傋傞丅偙偺曽朄偑儂儎偱傕偆傑偔偄偔偙偲偼macho-1偺働乕僗偱妋偐傔傜傟偰偄傞丅偙偺幚尡傪峴偄丄壓棳堚揱巕偺Brachyury偲actin偺敪尰傪挷傋傞偲丄Ets偑揮幨妶惈壔場巕側偺偐丄偦傟偲傕梷惂壔場巕側偺偐丄偼偨傑偨壓棳堚揱巕偵埶懚偟偰偳偪傜偵傕側傟傞偐傪妋幚偵敾掕偱偒傞丅
丂Ets偲macho-1偼捈愙偵偼寢崌偟側偄偐傕偟傟側偄丅偦偺応崌偼丄偙傟傜偺忣曬偺摑崌偼丄壓棳堚揱巕偺敪尰惂屼椞堟忋偱峴傢傟傞偲峫偊傜傟傞丅巹偼埲慜偺尋媶偱丄僾儘儌乕僞乕夝愅傪偍偙側偭偰偒偨丅偙偺宱尡傪惗偐偟偰丄Ets傕偟偔偼macho-1偺偡偖壓棳偱塭嬁傪庴偗傞堚揱巕偺僾儘儌乕僞乕夝愅傪峴偆帠偵傛偭偰丄壓棳堚揱巕偺揮幨挷愡婡峔傪徻嵶偵挷傋偨偄丅Ets偲macho-1偺塭嬁傪捈愙庴偗偰偄傞偲梊憐偝傟傞堚揱巕偺僾儘儌乕僞乕傪暋悢僋儘乕僯儞僌偟偰揮幨挷愡椞堟傪夝愅偡傞丅桿摫偺寢壥丄梊掕愐嶕妱媴偱敪尰偡傞堚揱巕偲偟偰偼丄Brachyury偑偁傞丅娫廩怐偱macho-1偵傛傝妶惈壔偝傟Ets偵傛傝嵞傃梷惂偝傟傞堚揱巕偲偟偰偼傾僋僠儞丄儈僆僔儞丄Ca僩儔儞僗億乕僞乕丄Tbx-6側偳偑偁傞丅偙傟傜偺堚揱巕嬤朤偱Ets
寢崌攝楍傪専嶕偡傞偲偲傕偵丄儂儎棏傊堚揱巕摫擖偟丄Ets偲macho-1墳摎攝楍傪儗億乕僞乕傾僢僙僀偵傛傝僗僋儕乕僯儞僌偡傞丅偦偺屻Ets偲macho-1偺寢崌偡傞攝楍傪僎儖僔僼僩偵傛傝摿掕偡傞丅
丂偙偺傛偆偵Ets偲macho-1偺椉揮幨場巕偑偳偺傛偆偵嫤挷揑偵摥偔偐偵偮偄偰夝愅偡傞偲偲傕偵丄桿摫尰徾偵偍偗傞嵶朎撪場巕偍傛傃嵶朎奜僔僌僫儖偺摑崌偵娭偟偰摯嶡傪峴偆丅
擭師寁夋
- 侾擭栚
丒Ets偲macho-1抈敀幙偺捈愙偺憡屳嶌梡偵偮偄偰挷傋傞丅戝挵嬠敪尰儀僋僞乕偵Ets偲macho-1傪慻傒崬傒丄戝挵嬠偵敪尰偝偣惛惢偡傞丅嶌惉偟偨儕僐儞價僫儞僩僞儞僷僋幙傪梡偄偰僾儖僟僂儞傾僢僙僀朄傪梡偄偨寢崌幚尡傪峴偆丅昁梫偱偁傟偽Ets僞儞僷僋幙傪僉僫乕僛偱儕儞巁壔偟丄偦傟傕梡偄偰傒傞丅
丒峈懱偵傛傞僞儞僷僋幙偺敪尰応強偲妶惈壔応強偺摿掕丅Ets丄儕儞巁壔Ets丄macho-1偺儕僐儞價僫儞僩僞儞僷僋幙傪梡偄偰丄峈懱偺嶌惉傪峴偆丅嶌惉偟偨峈懱傪梡偄偰偄傠偄傠側敪惗僗僥乕僕偵偍偗傞僞儞僷僋偺敪尰応強傪摿掕偡傞丅
俀擭栚
丒儂儎偺Ets偑揮幨梷惂宆偐妶惈壔宆偐丄傑偨偼偦偺椉曽偺揮幨場巕偲偟偰摥偔偺偐傪挷傋傞丅Ets偲揮幨妶惈壔僪儊僀儞VP16傕偟偔偼梷惂壔僪儊僀儞EnR傪梈崌偝偣偨mRNA傪嶌惉偟丄儂儎泱偵尠旝拲擖偡傞丅宍懺傗暘壔儅乕僇乕偱嵶朎偺暘壔忬懺傪僠僃僢僋偡傞丅
丒壓棳堚揱巕偺敪尰惂屼椞堟偺僋儘乕僯儞僌丅Ets丄macho-1偺壓棳偱摥偔暋悢偺堚揱巕偺僾儘儌乕僞乕傪僎僲儉偐傜僋儘乕僯儞僌偡傞丅攝楍偺夝愅傪峴偭偨屻丄偙傟傜偺僾儘儌乕僞乕傪儖僔僼僃儔乕僛儗億乕僞乕偺儀僋僞乕偵慻傒崬傓丅
俁擭栚
丒Ets丄macho-1偑僞乕僎僢僩堚揱巕偺揮幨傪僾儘儌乕僞乕忋偱偳偺傛偆偵惂屼偡傞偐傪挷傋傞丅FGF張棟丄傕偟偔偼Ets丄macho-1偺堎強揑敪尰偵墳摎偡傞僾儘儌乕僞乕傪僗僋儕乕僯儞僌偡傞丅偙傟傜偺僾儘儌乕僞乕偺僨儗乕僔儑儞僐儞僗僩儔僋僩傪嶌惉偟丄Ets偍傛傃macho-1偵墳摎偡傞攝楍傪尷掕偟偰偄偔丅Ets傕偟偔偼macho-1偑寢崌偡傞攝楍傪僎儖僔僼僩傾僢僙僀偵傛傝寛掕偡傞丅
偙偺尋媶偺摿怓偲撈憂揑側揰
- 丂摦暔偺敪惗偱偼丄條乆側桿摫嶌梡偑廳梫側摥偒傪偟偰偄傞偙偲偑抦傜傟偰偄傞丅偟偐偟丄嬤擭偺抦尒偵傛傞偲丄嵶朎娫僔僌僫儖偲偟偰摥偗傞暘巕偺庬椶偑尷傜傟偰偍傝丄摦暔偼摨偠僔僌僫儖暘巕傪條乆側帪娫揑丒嬻娫揑嬊柺偱巊偄夞偟偟偰偄傞偙偲偑帵偝傟偰偄傞丅偡側傢偪丄摨偠僔僌僫儖傪庴偗庢偭偨偲偟偰傕丄嵶朎偼偦傟偧傟偺忬嫷偵墳偠偰堎側傞墳摎傪帵偡丅偙偺偙偲偼丄嵶朎偺墳摎擻偺惂屼偑偙傟偐傜偺桿摫尰徾偺夝愅偵偍偄偰傑偡傑偡廳梫帇偝傟傞偱偁傠偆偙偲傪梊應偝偣傞丅偙偺尋媶偱偼丄桿摫尰徾偵偍偄偰丄嵶朎奜偐傜嵶朎撪偵揱偊傜傟傞僔僌僫儖偐傜偺忣曬偲丄嵶朎撪偵偁偭偰嵶朎偺墳摎擻傪巟攝偟偰偄傞場巕偐傜偺忣曬偑丄偄偐偵嵶朎撪偱摑崌偝傟傞偐傪柧傜偐偵偡傞偙偲傪栚揑偲偟偰偄傞丅偙偙偱夝愅偟傛偆偲偟偰偄傞宯偱偼丄嵶朎奜僔僌僫儖偼FGF偱偁傝丄嵟廔揑偵Ets揮幨場巕偺妶惈壔傪傕偨傜偡丅斀墳擻傪寛傔偰偄傞嵶朎撪場巕偼macho-1揮幨場巕偱偁傞偙偲偑傢偐偭偰偄傞丅偙偺宯偼丄偦偺僔儞僾儖偝偲丄偡偱偵僉乕偲側傞暘巕偑傢偐偭偰偄傞偲偄偆偙偲偵偍偄偰懡戝側棙揰傪帩偭偰偍傝丄桿摫尰徾偺奣擮宍惉忋廳梫側峷專傪傕偨傜偡偙偲偑婜懸偝傟傞丅
丂杮尋媶偱偼丄愐嶕摦暔偱偁傞儂儎偺泱偲偄偆僔儞僾儖側僔僗僥儉傪夝愅懳徾偲偟偰偄傞丅10寧偐傜5寧偵偐偗偰偺僔乕僘儞拞偼丄枅擔嶻棏偝偣庴惛偝偣傞偙偲偑偱偒傞偺偱丄旕忢偵憗偄僒僀僋儖偱幚尡偑峴偊傞丅傑偨丄庴惛偐傜桿摫偑婲偙傞帪婜傑偱偼6帪娫偱偁傞丅偦偟偰丄嵶朎宯晥偑徻嵶偵傢偐偭偰偍傝丄妱媴侾偮侾偮偺敪惗塣柦偺夝愅偑壜擻偱偁傞丅泱憖嶌偑峴偊傞偨傔丄嵶朎偺暘壔傪恖堊揑偵憖嶌偟側偑傜丄傂偲偮傂偲偮偺嵶朎儗儀儖偱夝愅傪峴偆偙偲偑偱偒傞偺偼丄懠偱偼椶傪尒側偄摿挜偱偁傞丅摿偵強懏婓朷愭偺惣揷尋媶幒偼丄偙偺庬偺幚尡媄弍偵弉払偟偰偍傝丄徻嵶側夝愅曽朄偲丄僋儕傾乕側幚尡寢壥傪摼傞偨傔偺曽朄榑偵廏偱偰偄傞偲巚偭偰偄傞丅桿摫尰徾偵娭傢傞暘巕揑側婡峔傪夝愅偡傞偺偵傕丄偙傟傜偺儂儎泱偺摿挜偼戝偒偔峷專偡傞偩傠偆丅
丂傑偨丄偙傟傑偱偺夝愅偵傛傝偡偱偵僉乕偲側傞暘巕偑敾柧偟偰偄傞偲偄偆偙偲傕丄崱夞偺僥乕儅偵娭偡傞夝愅傪峴偆偵偁偨傝丄戝偒側棙揰偱偁傠偆丅尋媶傪巒傔傞偵摉偨傝丄弨旛偑廫暘偵惍偭偰偄傞偲偄偆偙偲傪堄枴偟偰偄傞丅偍偦傜偔丄杮尋媶偵傛傝僥乕儅偵増偭偨桳梡側寢壥偑摼傜傟傞壜擻惈偼偐側傝崅偄偲巚傢傟傞丅尋媶懳徾偵偟偰偄傞俀偮偺揮幨場巕偼嫽枴怺偄摿挜傪帩偭偰偄傞丅FGF僔僌僫儖偼丄偝傑偞傑側慻怐傊偺桿摫擻傪帩偭偰偄傞偙偲偑抦傜傟偰偍傝丄儂儎偵偍偄偰偼恄宱嵶朎傗愐嶕慻怐丄娫廩怐慻怐傊偺桿摫擻傪帩偮丅偳偺嵶朎僞僀僾偑桿摫偝傟傞偐偼僔僌僫儖傪庴偗庢傞懁偺嵶朎偵埶懚偟偰偄傞丅FGF偵傛傝妶惈壔偝傟傞Ets偼儂儎偺傒側傜偢丄條乆側嵶朎娫憡屳嶌梡偱巊傢傟偰偄傞偙偲偑傢偐偭偰偄傞丅堦曽丄傕偆侾偮偺場巕macho-1偼嬊嵼惈傪帵偡曣惈堚揱巕偱偁傞丅macho-1偼丄嵶朎偺墳摎擻傪僐儞僩儘乕儖偟偰偍傝丄摿堎揑側暘巕偱偁傝丄堦斒揑側Ets僔僌僫儖偵懳偡傞嵶朎偺墳摎傪摿堎壔偟偰偄傞丅嵶朎奜偲嵶朎撪偺忣曬偼丄偙偺傛偆偵揮幨場巕傪捠偟偰摑崌偝傟傞偙偲偑傢偐偭偰偄傞偑丄尰嵼偺強偦偺屻偵壗偑婲偙傞偐偑僽儔僢僋儃僢僋僗偲側偭偰偄傞丅
丂偙偺尋媶偱偼丄椉揮幨場巕偺捈愙偺憡屳嶌梡偺壜擻惈丄傕偟偔偼丄壓棳堚揱巕偺敪尰傪偦偺惂屼椞堟傪捠偟偰屳偄偵撈棫偵惂屼偟偰偄傞壜擻惈偺椉曽偵偮偄偰専摙偟丄桿摫尰徾偺堦斒揑側棟夝偵偮側偑傞偙偲傪婜懸偟偰丄尋媶傪寁夋偟偨丅偙偺傛偆偵丄侾丏偁傑傝夝愅偝傟偰偙側偐偭偨偑偙傟偐傜廳梫偵側傞偱偁傠偆丄嵶朎奜僔僌僫儖偲嵶朎撪場巕偺忣曬偺摑崌傪夝愅偡傞偙偲丄傑偨丄俀丏儂儎偲偄偆僔儞僾儖偱偐偮懡偔偺棙揰傪帩偭偨宯傪梡偄傞偙偲丄俁丏偙偺僥乕儅偵増偭偨幚尡傪峴偆偨傔偵婡偑弉偟偰偄傞偙偲側偳傪丄偙偺尋媶偺摿怓丒撈憂揑側揰偲偟偰嫇偘傞偙偲偑偱偒傞丅
妛弍嶨帍摍偱敪昞偟偨榑暥
- 徏杮弫乮拀攇戝妛堛妛尋媶壢攷巑壽掱乯丄拞杮愮徎乮崅抦戝棟妛尋媶壢戝妛堾乯丄摗尨帬庽乮崅抦戝棟妛晹彆嫵庼乯丄巜媧弐師乮崅抦戝棟妛晹嫵庼乯丄愳懞榓晇乮崅抦戝棟妛晹嫵庼乯
A
novel C-type lectin regulating cell growth, cell adhesion and cell
differentiation of the multipotent epithelium in budding
tunicates
Development 2001 Sep; 128(17): 3339-47
- 彑嶳桾1乮億僗僪僋乯丄徏杮弫乮拀攇戝妛堛妛尋媶壢攷巑壽掱乯1丄
壀揷弐徍乮億僗僪僋乯丄戝捤岾梇乮嶻憤尋尋媶堳乯丄捖楁乮億僗僪僋乯丄壀屗惏惗乮搶嫗搒恄宱尋庡擟尋媶堳乯丄壀懞峃巌乮嶻憤尋庡擟尋媶堳丄壀嶈崙棫尋媶強嫵庼暪擟乯
1Y.K.
and J.M. contributed equally to this work
Regulation of Synaptotagmin Gene Expression
during Ascidian Embryogenesis
Developmental Biology 2002 April;
244,293-304
妛夛摍偱敪昞偟偨榑暥
- 戝怷徣屷丄徏杮弫丄摗尨帬庽丄愳懞榓晇丄巜媧弐師
儈僒僉儅儊僀僞儃儎偺暘壔懡擻惈姴嵶朎偺嬅廤偵娭梌偡傞儗僋僠儞偺敪尰偲婡擻偺夝愅
擔杮摦暔妛夛戞俇俈夞戝夛丄嶥杫丄侾俋俋俇擭俋寧侾俉擔乕俀侽擔
徏杮弫丄戝怷徣屷丄摗尨帬庽丄愳懞榓晇丄巜媧弐師
儈僒僉儅儊僀僞儃儎偐傜扨棧偝傟偨怴婯儗僋僠儞偺峔憿偍傛傃婡擻夝愅
擔杮摦暔妛夛戞俇俉夞戝夛丄撧椙丄侾俋俋俈擭侾侽寧俀擔乕係擔
- 徏杮弫丄戝怷徣屷丄摗尨帬庽丄愳懞榓晇丄巜媧弐師
儈僒僉儅儊僀僞儃儎儗僋僠儞TC-30偺峔憿偲婡擻夝愅
戞31夞擔杮敪惗惗暔妛夛丄孎杮丄侾俋俋俉擭俆寧俀俋擔
- 徏杮弫丄摗尨帬庽丄愳懞榓晇丄巜媧弐師
弌夎儂儎儗僋僠儞偵傛傞忋旂惈嵶朎傊偺暘壔桿摫
戞俁俀夞擔杮敪惗惗暔妛夛丄恄屗丄1999擭俆寧俀俉乕俁侽擔
- 徏杮弫丄彑嶳桾丄壀懞峃巌
儂儎僔僫僾僩僞僌儈儞堚揱巕偵偍偗傞僯儏乕儘儞摿堎揑側敪尰偵娭傢傞僔僗僄儗儊儞僩偺夝愅
戞俁俁夞擔杮敪惗惗暔妛夛丄崅抦丄俀侽侽侽擭俆寧俀俆乕俀俈擔
- Jun Matsumoto,丂You Katsuyama and Yasushi Okamura
Multiple cis-regulatory regions control neuronal gene expression
of synaptotagmin in ascidian embryos
The First International
Symposium on the Biology of Ascidians乮Sapporo, Japan乯 June
26-30,2000
丂丂偙偺妛夛偺撪梕偼埲壓偺僾儘僔乕僨傿儞僌偱敪昞
丂丂丂丂J. Matsumoto, Y. Katsuyama, Y.
Ohtsuka and Y. Okamura
丂丂丂丂The Biology of Ascidian (Springer):
158-161
丂丂丂丂Multiple cis-regulatory regions control neuronal gene
expression of synaptotagmin in ascidian embryos
- J. Matsumoto, Y. Katsuyama, Y. Ohtsuka and Y. Okamura
Multiple cis-regulatory regions control neuronal gene expression
of synaptotagmin in ascidian embryos
14th international congress of
developmental biology (Kyoto, Japan) July 8-12,2001
- 徏杮弫丄壀懞峃巌
俀庬偺儂儎傪梡偄偨僔僫僾僩僞僌儈儞堚揱巕偺僾儘儌乕僞乕夝愅
戞俁俆夞擔杮敪惗惗暔妛夛丄墶昹丄俀侽侽俀擭俆寧俀侾亅俀俁擔
傕偭偲傕庡梫側榑暥偺梫巪
- Development 128, 3339-3347 (2001)
A novel C-type
lectin regulating cell growth, cell adhesion and cell differentiation of the
multipotent epithelium in budding
tunicates
乮儈僒僉儅儊僀僞儃儎偵偍偗傞暘壔懡擻惈忋旂嵶朎偺嵶朎憹怋丄嵶朎愙拝丄嵶朎暘壔傪挷愡偡傞怴婯C宆儗僋僠儞乯
J.
Matsumoto, C. Nakamoto, S. Fujiwara, T. Yubisui, and K.
Kawamura
丂柍惈惗怋傗嵞惗偺夁掱偵偍偄偰丄屄懱偺慻怐傗婍姱偺嵞峔抸偵偼丄枹暘壔側姴嵶朎偺憹怋偲暘壔偑昁梫偱偁傞丅尨嶕摦暔偺儈僒僉儅儊僀僞儃儎偼弌夎偲偄偆柍惈惗怋偵傛傝丄怴偨側屄懱傪宍惉偡傞丅偙偺宯偵偍偄偰丄姴嵶朎偺栶妱傪扴偆偺偑埻橐峯忋旂偲屇偽傟傞暘壔懡擻惈傪帩偮嵶朎偱偁傞丅弌夎夁掱偵偍偄偰偼丄偙偺嵶朎偑偄偭偨傫扙暘壔偡傞偙偲偵傛傝丄徚壔娗傗橐側偳偺婍姱嵞暘壔偟偰宍懺宍惉偑峴傢傟傞偲峫偊傜傟偰偄傞丅傑偨丄偙偺暘壔懡擻惈偺忋旂嵶朎傪攟梴偡傞偲丄埻橐峯忋旂偲偟偰偺暘壔忬懺傪幐偄丄憹怋傪懕偗傞偙偲偑傢偐偭偰偄傞丅偦偺惈幙傪棙梡偟偰埻橐峯忋旂儂儎偺攟梴嵶朎姅偑愳懞傜偵傛偭偰妋棫偝傟丄宲戙攟梴偝傟偰偄傞丅偙偺儂儎攟梴嵶朎偵懳偟偰丄嵶朎嬅廤傗嵶朎憹怋梷惂嶌梡傪帩偮惗棟妶惈場巕偑儈僒僉儅儊僀僞儃儎帺懱偺儂儌僕僱乕僩拪弌塼拞偵敪尒偝傟偨丅
丂偙偺榑暥偱偼偙偺惗棟妶惈場巕偺摨掕偍傛傃夝愅傪峴偭偰偍傝丄巹偼埲壓偺幚尡傪扴摉偟偨丅傑偢丄儈僒僉儅儊僀僞儃儎儂儌僕僱乕僩偐傜丄儂儎攟梴嵶朎偵懳偡傞嵶朎嬅廤妶惈傪巜昗偵偟偰丄偙偺惗棟妶惈場巕偺惛惢傪恑傔偨丅僎儖傠夁僋儘儅僩僌儔僼傿乕丄堿僀僆儞岎姺僋儘儅僩僌儔僼傿乕丄僎儖傠夁HPLC偲惛惢傪恑傔偨偲偙傠丄18kDa偲15kDa偺儁僾僠僪偑扨棧偝傟偨丅偙偺俀偮偺億儕儁僾僠僪乮P18/P15乯偼SDS-PAGE忋偱傎傏1:1偺検斾偱懚嵼偟偰偍傝丄僋儘儅僩僌儔僼傿乕偲僎儖傠夁HPLC偱偼奺乆傪暘棧偡傞偙偲偼偱偒側偐偭偨丅偦偙偱丄P18/P15偺傾儈僲巁攝楍傪晹暘揑偵僔乕僋僄儞僗偟偨丅偡傞偲Blast僒乕僠偵傛傝丄埲慜偵儈僒僉儅儊僀僞儃儎偱尒偮偐偭偨僈儔僋僩乕僗寢崌C宆儗僋僠儞TC14-1(tunicate
C-type lectin
TC14-1)偵崅偄椶帡惈偑偁傞偙偲偑傢偐偭偨丅師偵丄P18/P15偺摐寢崌擻傪僈儔僋僩乕僗屌掕壔傾僼傿僯僥傿乕僇儔儉偱挷傋偨丅P18/P15偼僇儖僔僂儉懚嵼壓偱僈儔僋僩乕僗屌掕壔傾僼傿僯僥傿乕僇儔儉偵寢崌偟丄僇儖僔僂儉傪EDTA偱僉儗乕僩偡傞偙偲偱梟弌偟偰偒偨丅偦偟偰丄梟弌偟偨P18/P15偼儂儎攟梴嵶朎偵懳偡傞嵶朎嬅廤妶惈傪帩偭偰偄偨丅偮傑傝丄P18/P15偼儂儎攟梴嵶朎偵懳偡傞嵶朎嬅廤妶惈傪帩偮儗僋僠儞偱偁傞偙偲偑妋掕偟偨丅師偵丄晹暘傾儈僲巁攝楍傪傕偲偵慡挿cDNA傪僋儘乕僯儞僌偟偨丅P18偼埲慜偵cDNA抐曅偑曬崘偝傟偰偄偨TC14-2偲姰慡偵堦抳偟偰偄偨丅偦偟偰丄P15偼怴婯偵尒偮偐偭偨偺偱TC14-3偲柤偯偗偨丅偙傟傜俁偮偺儗僋僠儞偺梊憐傾儈僲巁攝楍偼偲傕偵145巆婎偱丄C宆儗僋僠儞偵摿挜揑側俉傾儈僲巁偑曐懚偝傟偰偍傝丄N枛抂偵偼20巆婎偺僔僌僫儖儁僾僠僪偲峫偊傜傟傞攝楍偑偁偭偨丅梊憐傾儈僲巁攝楍傪斾妑偡傞偲丄TC14-1偲TC14-2偼84.8%丄TC14-1偲TC14-3偼65.7%丄
TC14-2偲TC14-3偼65.0%偺椶帡惈傪帵偟偨丅
丂惗懱偐傜惛惢偟偨TC14-2/TC14-3乮P18/15乯偼僇儔儉惛惢偱屳偄偵暘棧偡傞帠偑偱偒側偐偭偨丅偦偙偱丄奺乆偺cDNA傪戝挵嬠敪尰儀僋僞乕偵慻傒崬傒儕僐儞價僫儞僩僞儞僷僋幙(rTC14-2丄rTC14-3)傪嶌惢偟偨丅rTC14-2偲rTC14-3偼丄偦傟偧傟扨撈偱僇儖僔僂儉埶懚揑偵僈儔僋僩乕僗偵寢崌偟偨丅偩偑丄儂儎攟梴嵶朎偵懳偟偰偼rTC14-3偺傒偑嵶朎嬅廤妶惈傪帵偟丄TC14-1偲rTC14-2偼嵶朎嬅廤妶惈傪帵偝側偐偭偨丅師偵丄rTC14-3偺帩偮僈儔僋僩乕僗寢崌擻偑儂儎攟梴嵶朎偵懳偡傞嵶朎嬅廤妶惈偵昁梫側偺偐挷傋偨丅僼儕乕偺僈儔僋僩乕僗傪TC14-3偲嫟偵嵶朎攟梴塼偵揧壛偟偰丄TC14-3偺僈儔僋僩乕僗寢崌僪儊僀儞傪僽儘僢僋偟偨丅偡傞偲丄50mM偺僈儔僋僩乕僗揧壛偱TC14-3偺嵶朎嬅廤妶惈偼姰慡偵慾奞偝傟偨丅偙偺寢壥偐傜丄TC14-3偺僈儔僋僩乕僗寢崌擻偼丄儂儎攟梴嵶朎偵懳偡傞嵶朎嬅廤妶惈偵昁恵偱偁傞帠偑帵偝傟偨丅側偤TC14-3偺傒偑儂儎攟梴嵶朎偵懳偡傞嵶朎嬅廤妶惈擻傪帩偮偺偐偼晄柧偱偁傞丅偦偺棟桼偺侾偮偲偟偰峫偊傜傟傞偺偼丄TC儗僋僠儞偺擣幆偡傞嵶朎昞柺偺摐嵔偺堘偄偱偁傞丅堦斒揑偵嵶朎昞柺偼僈儔僋僩乕僗偺傛偆側扨摐偱偼側偔懡摐偵側偭偰偍傝丄偦偺棫懱峔憿偼嵶朎昞柺偺忣曬偲偟偰棙梡偝傟偰偄傞丅崱夞幚尡偵梡偄偨埻橐峯忋旂桼棃偺儂儎攟梴嵶朎昞柺偺摐嵔忣曬偼TC14-3偺傒偑擣幆壜擻偱偁傝丄偦偺寢壥偲偟偰嵶朎嬅廤妶惈傪帵偟偨偲悇嶡偝傟偨丅
丂埻橐峯忋旂桼棃偺儂儎攟梴嵶朎偼丄宲戙攟梴壓偱偼扙暘壔偟偰偄傞偲峫偊傜傟偰偍傝丄埻橐峯忋旂偲偟偰偺暘壔忬懺傪帵偝側偄丅偦偙偱丄偙偺傛偆側惈幙偺嵶朎姅偵TC儗僋僠儞傪嶌梡偝偣偨帪偺暘壔忬懺傪挷傋偨丅暘壔偺巜昗偲偟偰丄埻橐峯忋旂傕偟偔偼徚壔娗偵懳偡傞摿堎揑峈懱乮偦傟偧傟AP-E偲AP-G乯傪梡偄偰峈懱愼怓傪峴偭偨丅TC14-1偲rTC14-2傪嶌梡偝偣偨嵶朎偼偳偪傜偺峈懱偵傕斀墳偟側偐偭偨丅偟偐偟丄rTC14-3傪嶌梡偝偣偨嵶朎偼丄AP-G偵偼斀墳偟側偐偭偨偑丄AP-E偵偼偡傋偰偺嵶朎偑嫮偔斀墳偟偨丅偮傑傝丄TC14-3偺嶌梡偵傛傝丄儂儎攟梴嵶朎偼傕偲偺埻橐峯忋旂偺暘壔忬懺偵桿摫偝傟偨偲峫偊傜傟傞丅
丂埲屻丄愳懞傜偵傛偭偰埲壓偺幚尡偑曗懌偝傟偨丅TC14-3偺嶌梡偵傛傞儂儎攟梴嵶朎偺嵶朎悢偺曄壔傪挷傋偨偲偙傠丄嵶朎憹怋偑梷惂偝傟傞偙偲偑傢偐偭偨丅傑偨丄TC14-3偺嶌梡偵傛傝嵶朎愙拝偵娭傢傞兛僀儞僥僌儕儞偑攟梴嵶朎昞柺偵桿摫偝傟傞偙偲偑傢偐偭偨丅兛僀儞僥僌儕儞偼儈僒僉儅儊僀僞儃儎惗懱撪偱偼埻橐峯忋旂嵶朎偱敪尰偟偰偄傞偺偱丄TC14-3偺嶌梡偵傛偭偰嵶朎愙拝偑桿摫偝傟丄攟梴嵶朎偼忋旂嵶朎偺宍懺偵側偭偰偄傞帠偑帵嵈偝傟偨丅TC14-3偺儌僲僋儘乕僫儖峈懱傪嶌惉偟僞儞僷僋偺敪尰応強傪挷傋偨偲偙傠丄恊屄懱偱偼埻橐峯忋旂偲寣媴嵶朎偱敪尰偑傒傜傟偨丅偙傟偵傛傝丄娫廩怐撪偺棳摦揑側寣媴嵶朎偺偆偪丄摿掕偺嵶朎偑TC14-3偺嶌梡傪庴偗丄埻橐峯忋旂偲偟偰暘壔偡傞偙偲偑帵嵈偝傟偨丅傑偨丄弌夎拞偺夎懱偱偼TC14-3偑摿偵嫮偔敪尰偟偰偍傝丄埻橐峯忋旂嵶朎偺嫙媼傪惙傫偵峴偭偰偄傞偲峫偊傜傟偨丅傑偨丄埻橐峯忋旂偑扙暘壔偡傞夎懱屻曽偺宍懺宍惉椞堟偱偼TC14-3偺敪尰偼徚幐偟偰偄偨丅傛偭偰丄宍懺宍惉偺僔僌僫儖傪庴偗偨椞堟偱TC14-3偑徚幐偡傞偙偲偑丄埻橐峯忋旂偺扙暘壔偵昁梫偩偲峫偊傜傟偨丅
丂埲忋丄偙偺榑暥偱偼丄怴婯偵僈儔僋僩乕僗寢崌C宆儗僋僠儞TC14-3傪僋儘乕僯儞僌偟丄儗僋僠儞偵傛傞嵶朎偺暘壔惂屼偵偮偄偰怴偨側曬崘傪峴偭偨丅